大黑星龜金花蟲的生殖行為與翅鞘顏色變化
組員:王靖云、李沁柔、曾姵榛
摘要
大黑星龜金花蟲(Aspidomorpha miliaris)為台灣常見的昆蟲之一,但相關文獻少有對於交配行為的描述。本研究分析記錄大黑星龜金花蟲七種交配行為模式:觸角移動、後肢抽動、身體搖動、振翅、輕抽身體、循環(後肢抽動加搖動身體的連續行為)、以及翅鞘變色,且它們之間存在著一定的連續關係。而交配時雌雄體型大小的差異會影響交配行為中雄蟲動作的頻率。針對可逆性的交配行為翅鞘變色及不可逆的翅鞘老化變色,我們利用掃描式電子顯微鏡觀察、比較翅鞘內部超顯微結構,探討翅鞘不可逆變色的結構因子,可能為針狀突起密度或薄層厚度的變化;而可逆性變色的機制,則可能為蠟質細絲或空腔內體液的分布改變。
研究動機
我們時常在校園圍牆的牽牛花植株上觀察到大黑星龜金花蟲的族群中,不同個體間會隨著羽化後日齡不同而擁有白、黃、橘三種主要的顏色差異;然而我們也注意到,其交配中的個體翅鞘顏色大多呈白色和淡黃色,引發我們的好奇,莫非大黑星龜金花蟲的翅鞘也會跟許多物種一樣利用變色來彰顯自己是理想的交配對象或增加繁殖成功率?而在變色時翅鞘內部又有什麼樣的細微變化呢?經過查詢文獻後發現相關研究十分缺乏,故決定研究其交配行為與翅鞘顏色變化之間的關聯性,探討交配展現變色等特定行為的原因及翅鞘變色機制。
研究目的
(一)生殖交配行為中展現的行為模式
(二)翅鞘中造成外部顏色變化的構造
(三)交配行為與翅鞘顏色變化之關聯
實驗方法
首先以錄影機紀錄影像中交配行為的出現時間、頻率,並對各項行為與雌雄蟲體型比例大小的關係進行分析;另以桌上型掃描式電子顯微鏡(SEM)觀察大黑星龜金花蟲翅鞘內部的構造。最後整合交配行為和翅鞘構造的觀察探討造成翅鞘變色的可能機制。
物種介紹
大黑星龜金花蟲 Aspidomorpha miliaris
(一)外型介紹
成蟲的翅鞘表面光滑無突起,在最外側展延成扁平裙狀。翅鞘依外觀顏色可分為身體區和透明區,表面皆有分布大小不同的斑點,故得其「大黑星」之中文俗名。成蟲性別的分辨方式在於雌蟲腹部的最末節具有一黑色條紋,雄蟲則無。
(二)日齡判斷
大黑星龜金花蟲成蟲的體色依羽化後日齡變化,呈白、黃、橘三種。根據觀察,成蟲翅鞘顏色變化的順序依序為剛羽化一小時內呈現的淡黃半透明(斑點尚未浮現)、羽化後14日內呈現的乳白色、羽化後60日內呈現的銘黃色(期間內有淺黃與深黃的細微變化)、羽化後60日以後呈現的深橘色。

實驗結果與討論
交配行為模式觀察
本研究藉由分次對於不同對交配中雌雄蟲所做的錄影紀錄,一共觀察並整理出七項雄蟲所具有的高辨識度行為模式,各說明如下:

各項交配行為之間的順序性
我們分析交配行為影片,記錄每一個動作接續的交配行為,並分別計算其機率。最後將每個行為後接續具有最高機率的動作,彙整出大黑星龜金花蟲的交配行為連續動作圖。
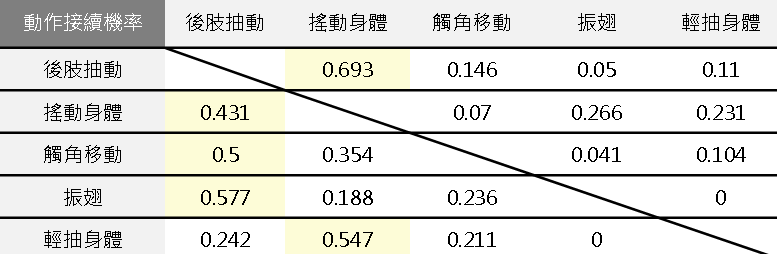
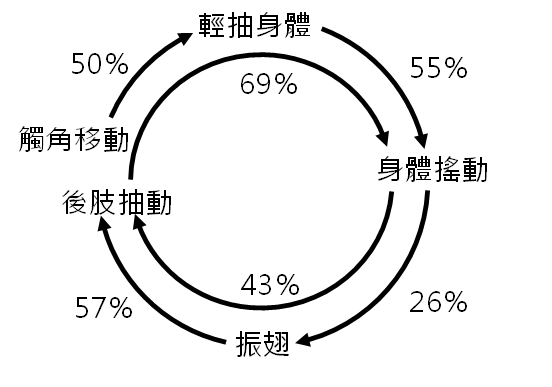
交配行為模式之意義分析結果
研究指出,龜金花蟲Acromis sparsa的交配行為在體型和武器(翅鞘尖端)較大或顯著的雄蟲身上明顯較少出現。因此觀察大黑星龜金花蟲的交配行為模式後,我們記錄了各行為的動作頻率,以及各對雌雄蟲體型大小比例差異,並加以進行分析。
(一)搖動身體頻率
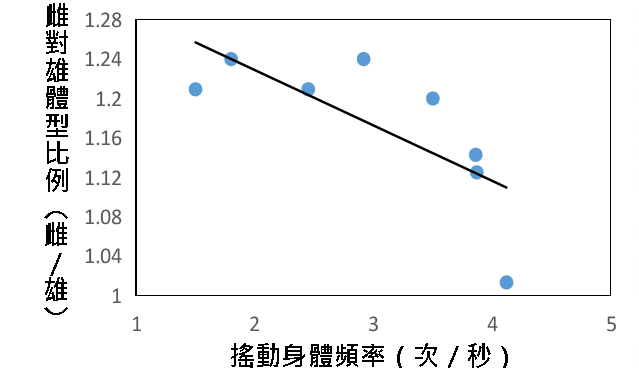
透過回歸分析可知,搖動身體頻率與雌雄體型呈中度負相關。雌雄體型差異越大,頻率越低。$$ R^{2} = 0.5387 $$
(二)後肢抽動頻率
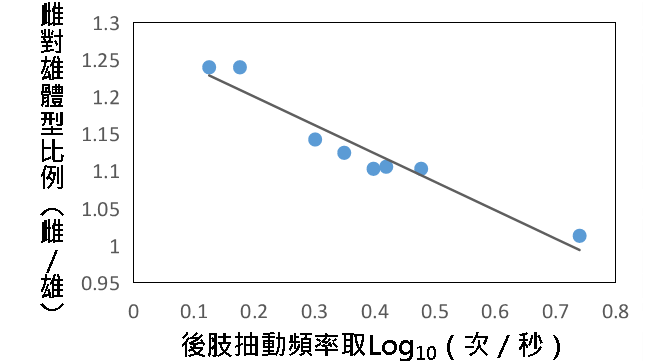
透過回歸分析可知,後肢抽動頻率與雌雄體型差異呈高度負相關。雌雄體型差異越大,頻率越低。$$ R^{2} = 0.9306 $$
(三)循環頻率
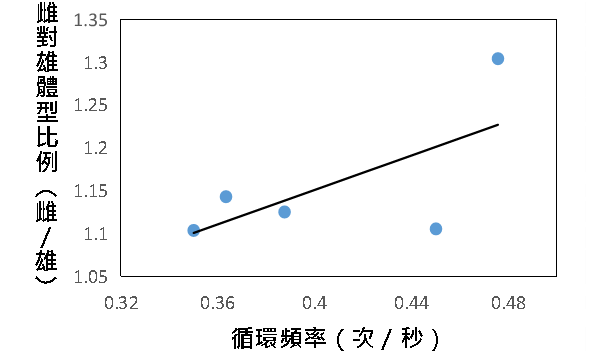
我們將大黑星龜金花蟲交配行為中反覆出現的一次循環定為:雄蟲後肢抽動 2 至 3 下再劇烈搖動身體。透過回歸分析可知,循環頻率與雌雄體型差異呈中度至低度正相關。雌雄體型差異越大,頻率越高。$$ R^{2} = 0.4305 $$
交配行為模式的生物意義
由以上結果顯示大黑星龜金花蟲交配時,雌雄蟲的體型差異確實會影響雄蟲表現之交配行為的動作頻率。參考論文後我們合理推測大黑星龜金花蟲的受精成功率和雌雄蟲體型差異大小有關,而部分雄蟲為了彌補未符合最佳受精成功率的體型差異之不利繁殖條件,利用了上述特殊的交配行為模式或改變交配行為的頻率作為提升受精成功率的手段。
翅鞘構造介紹
節肢動物發達的外骨骼,又稱表皮層,普遍由外而內可分為三個部分:上表皮(epicuticle)、外表皮(exocuticle)、與內表皮(endocuticle)。以下是我們透過掃描式電子顯微鏡SEM觀察所整理出的大黑星龜金花蟲翅鞘分層細部結構分佈。
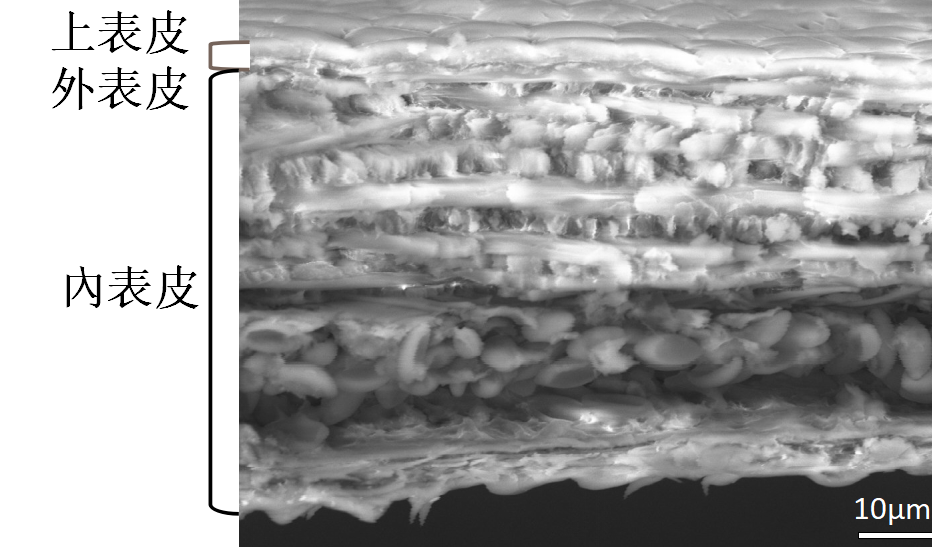
(一)薄層:佔內表皮比例最高,分為平面薄層(plies)和纖維薄層(balken)。平面薄層單一一層纖維又裂成更多平行的細小纖維,且從剖面能明顯看出相鄰兩層的纖維夾角約 90 度。但兩者厚度、性質、用途等,無太大差異。

(二)單凹橄欖:中空構造,分散生、叢生。單體為約 10μm×8.5μm 的單凹橄欖球型。散生者無規律的零星分布在薄層間;而叢生者則聚集在空腔內腹側,極似方便麵。
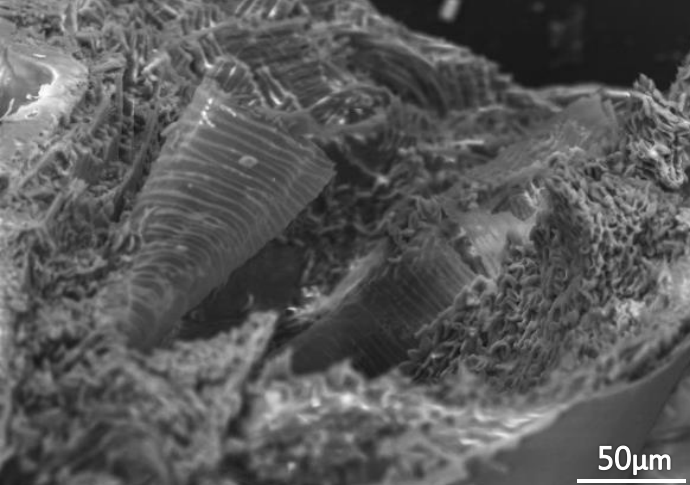
(三)空腔:體液流動處。分佈在內表皮中下層,薄層會以同心圓繞著空腔生長。

(四)針狀突起:空心結構,型態隨翅鞘部位有細微差異。

變色機構分析
(一) 自然老化的體色變化
1. 針狀突起的密度大小
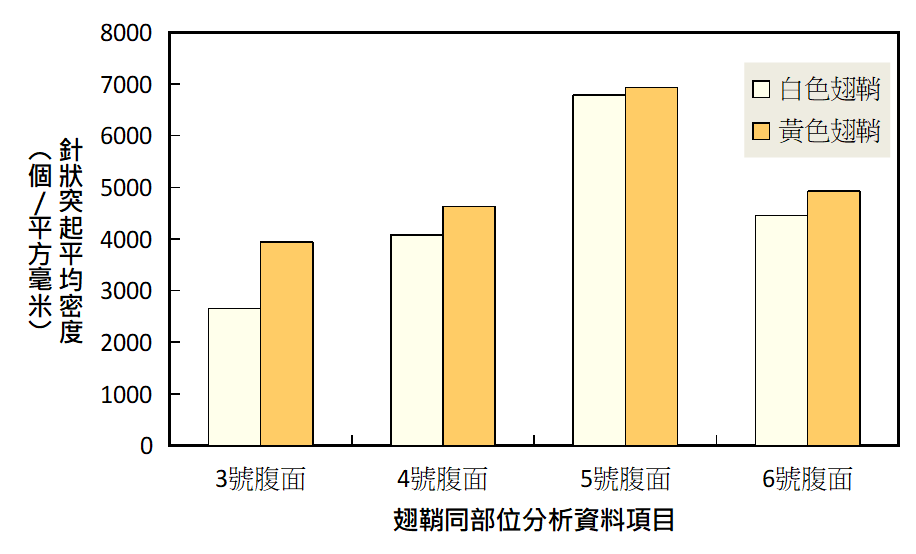
同一部位中黃色大於白色翅鞘的針狀突起平均密度。此現象與前人研究相符,針狀突起會隨著成蟲翅鞘的微結構增生而越來越多,可應用於判斷成蟲日齡。$$ p< 0.05 $$
2. 單一薄層的厚薄
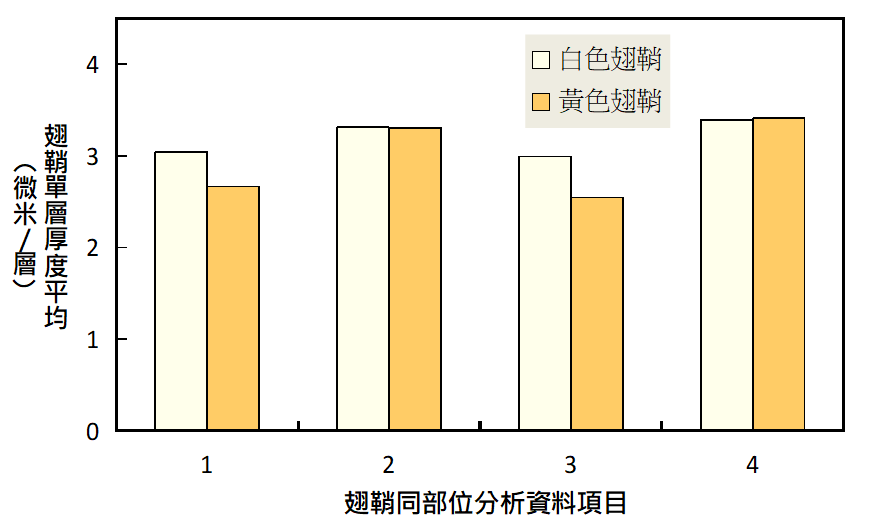
年輕成蟲的薄層平均較年老者厚。推測此現象為翅鞘中微結構差異造成,厚度改變影響了視覺上的顏色。
(二) 交配的體色變化
1. 細絲狀蠟的移動 (wax filament deposition/removal)
輸蠟通道(wax channel)貫穿上表皮至內表皮上層,使儲蠟空間中的蠟質細絲能覆蓋到上表皮,進而改變翅鞘的顏色。

2. 空腔內的體液多寡
以控制翅鞘中體液多寡改變體色的例子不罕見,且與大黑星龜金花蟲親緣關係接近的巴拿馬龜金花蟲Charidotella egregia也以此方式改變顯色。以此機制來變換顏色的機率仍頗高。

結論
一、大黑星龜金花蟲雄蟲具有高辨識度交配行為模式,分別為觸角移動、後肢抽動、身體搖動、振翅、輕抽身體、循環、以及翅鞘變色。且其交配行為具有特定順序。
二、大黑星龜金花蟲交配中搖動身體與後肢抽動的頻率與雌雄蟲體型差異有關。
三、翅鞘變色可分為老化造成的不可逆變色以及交配行為造成的可逆性變色。
四、翅鞘不可逆變色的微結構變化為針狀突起的密度或薄層厚度的變化;可逆性變色的機制推測為蠟質細絲或空腔內體液的分布改變。
參考資料
[1] Chen, J., Zu, Q., Wu, G., Xie, J., & Tuo, W. (2015). Review of beetle forewing structures and their biomimetic applications in China:(II) On the three-dimensional structure, modeling and imitation. Materials Science and Engineering: C, 55, 620-633.
[2] Du, J., & Hao, P. (2018). Investigation on Microstructure of Beetle Elytra and Energy Absorption Properties of Bio-Inspired Honeycomb Thin-Walled Structure under Axial Dynamic Crushing. Nanomaterials, 8(9), 667.
[3] Kundanati, L., Signetti, S., Gupta, H. S., Menegon, M., & Pugno, N. M. (2018). Multilayer stag beetle elytra perform better under external loading via non-symmetric bending properties. Journal of The Royal Society Interface, 15(144), 20180427.
[4] Sun, J., & Bhushan, B. (2012). Structure and mechanical properties of beetle wings: a review. Rsc Advances, 2(33), 12606-12623.
[5] Tang, Z. J., Lue, S. I., Tsai, M. J., Yu, T. L., Thiyagarajan, V., Lee, C. H., ... & Weng, C. F. (2014). The hormonal regulation of color changes in the sexually dichromatic frog Buergeria robusta. Physiological and Biochemical Zoology, 87(3), 397-410.
[6] Trillo, P. A. (2008). Pre-and post-copulatory sexual selection in the tortoise beetle Acromis sparsa (Coleoptera: Chrysomelidae). (Unpublished doctoral dissertation).The University of Montana.
[7] Umbers, K. D., Fabricant, S. A., Gawryszewski, F. M., Seago, A. E., & Herberstein, M. E. (2014). Reversible colour change in Arthropoda. Biological Reviews, 89(4), 820-848.
[8] Van de Kamp, T., & Greven, H. (2010). On the architecture of beetle elytra. Entomologie heute, 22, 191-204.
[9] Van de Kamp, T., Riedel, A., & Greven, H. (2016). Micromorphology of the elytral cuticle of beetles, with an emphasis on weevils (Coleoptera: Curculionoidea). Arthropod structure & development, 45(1), 14-22.
[10] Vigneron, J. P., Pasteels, J. M., Windsor, D. M., Vértesy, Z., Rassart, M., Seldrum, T., ... & Ertz, D. (2007). Switchable reflector in the Panamanian tortoise beetle Charidotella egregia (Chrysomelidae: Cassidinae). Physical Review E, 76(3), 031907.
[11] 李奇峰、鄭興宗(2007).台灣產金花蟲科圖誌Ⅰ.四獸山昆蟲相調查網。
[12] 鄭仲良, 楊正澤, & 蕭文鳳. (2015). 大黑星龜金花蟲 (Aspidomorpha miliaris (Fabricius 1775))(鞘翅目: 金花蟲科) 各期形態及翅鞘斑紋多型性描述. 台灣昆蟲, 35(1), 23-34.